







Master's Perspective
肝臓をめぐる旅
静岡大学 塩尻信義
はじめに
私の研究対象としての肝臓との出会いは、大学院時代の指導教授水野丈夫先生(現東大名誉教授)から与えられた研究テーマにさかのぼる。以来、肝臓との付き合いは46年ほどになるが、こんなに長い付き合いになるとは当時は思いもよらなかった。元々、高校の生物の授業で習ったシュペーマンのオーガナイザーについて感動し、大学では発生学を是非学びたいと思っていた。研究を進める中、成果が思ったように出ず、肝臓以外のテーマを考えたこともあった。今はこの46年を素直に驚くとともに、肝細胞研究会を含め色々な人との出会いが私の研究のターニングポイントでの大きな鍵になっていたり、また貴重なサポートでもあったと考えている。私の研究成果は中途半端なものが多く、今のシャープな研究アプローチには馴染まず、そのため若いみなさんの参考にならないのではと思いつつも、ここでは私の肝臓研究(“肝臓をめぐる旅”)のはじまりについて紹介したい。肝臓の世界はまだまだ未開拓なところが多く、この小文が興味のある方の参考になるとしたら望外の喜びである。
マウスにおける肝外および肝内胆管の発生
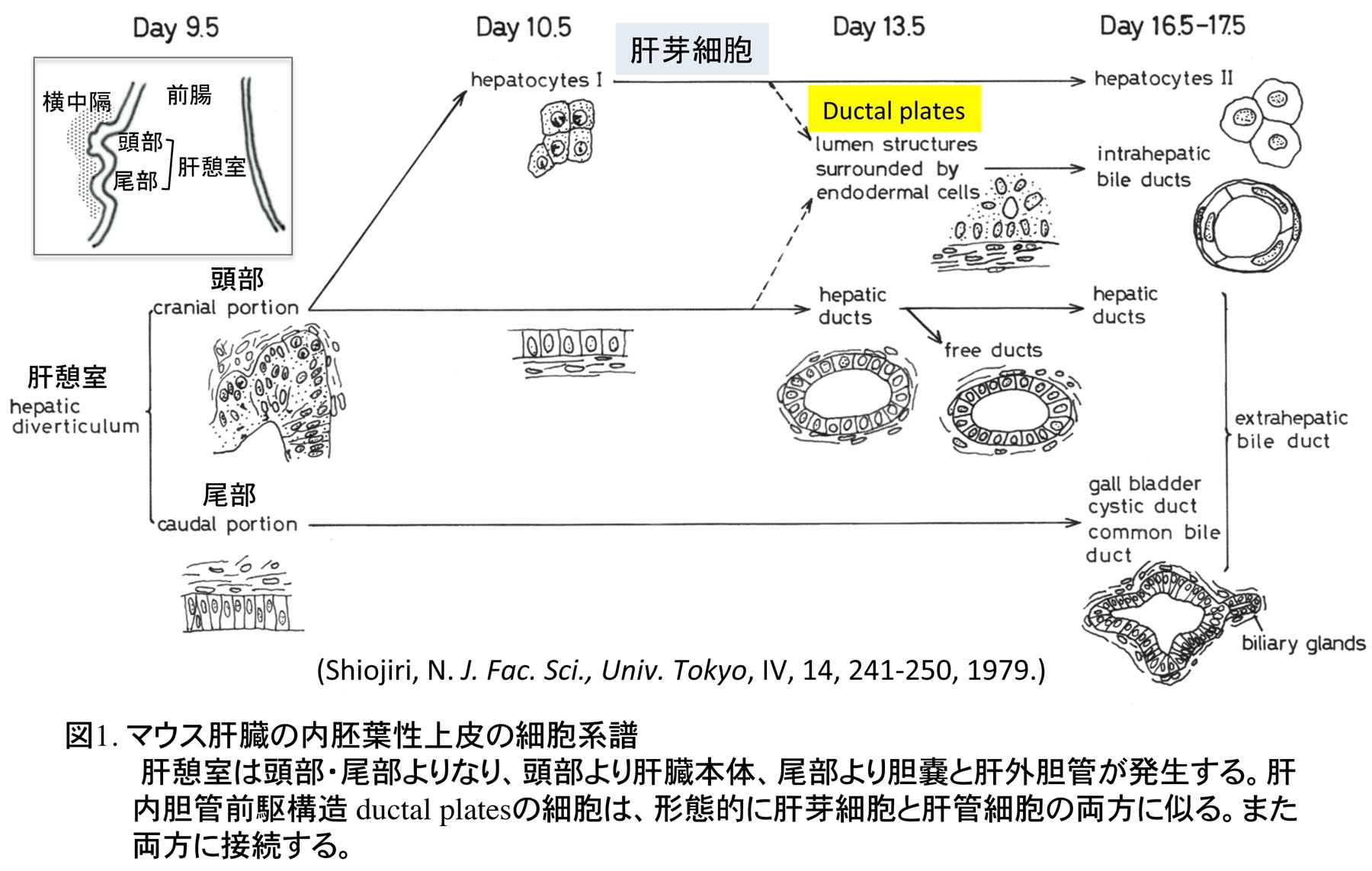
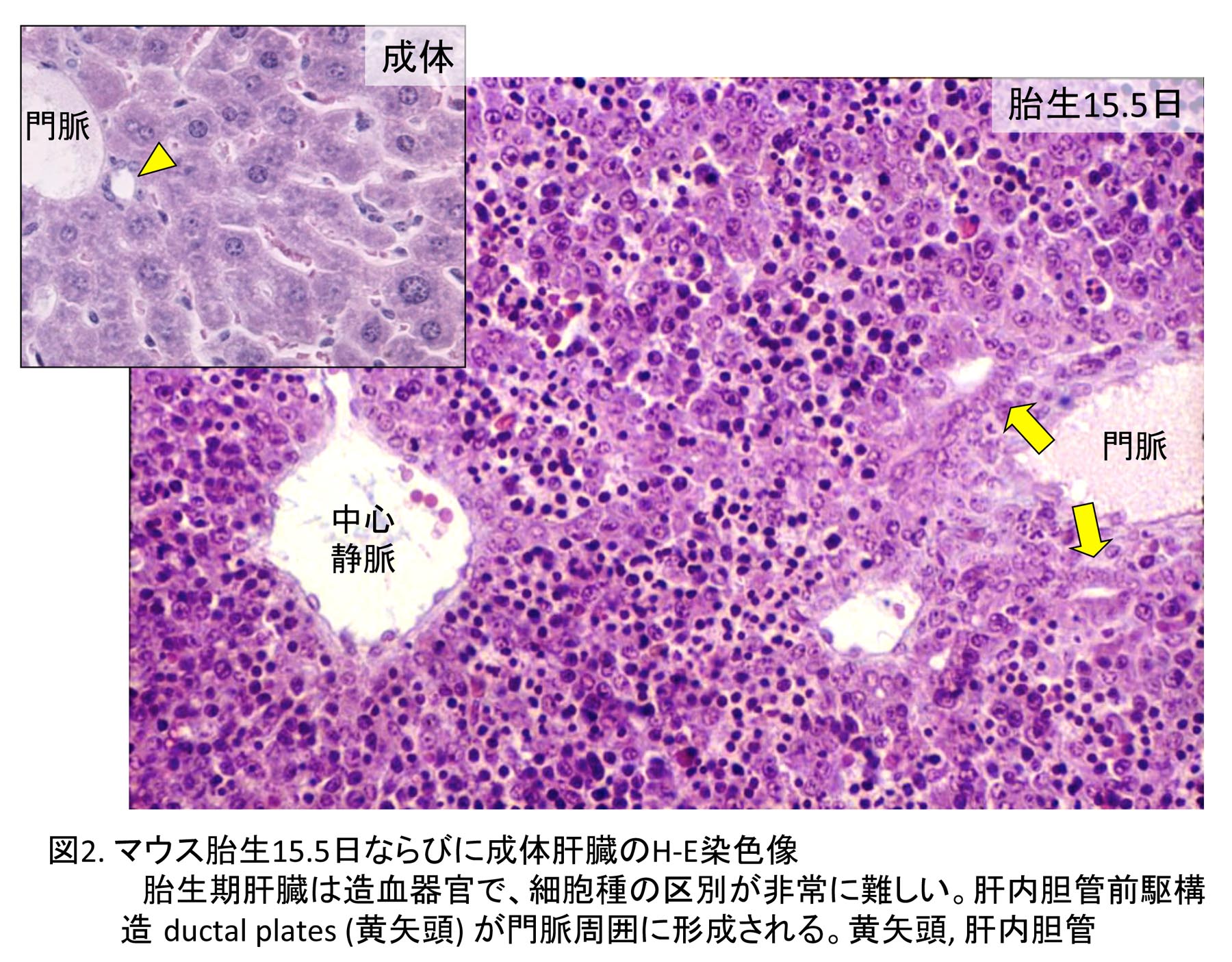
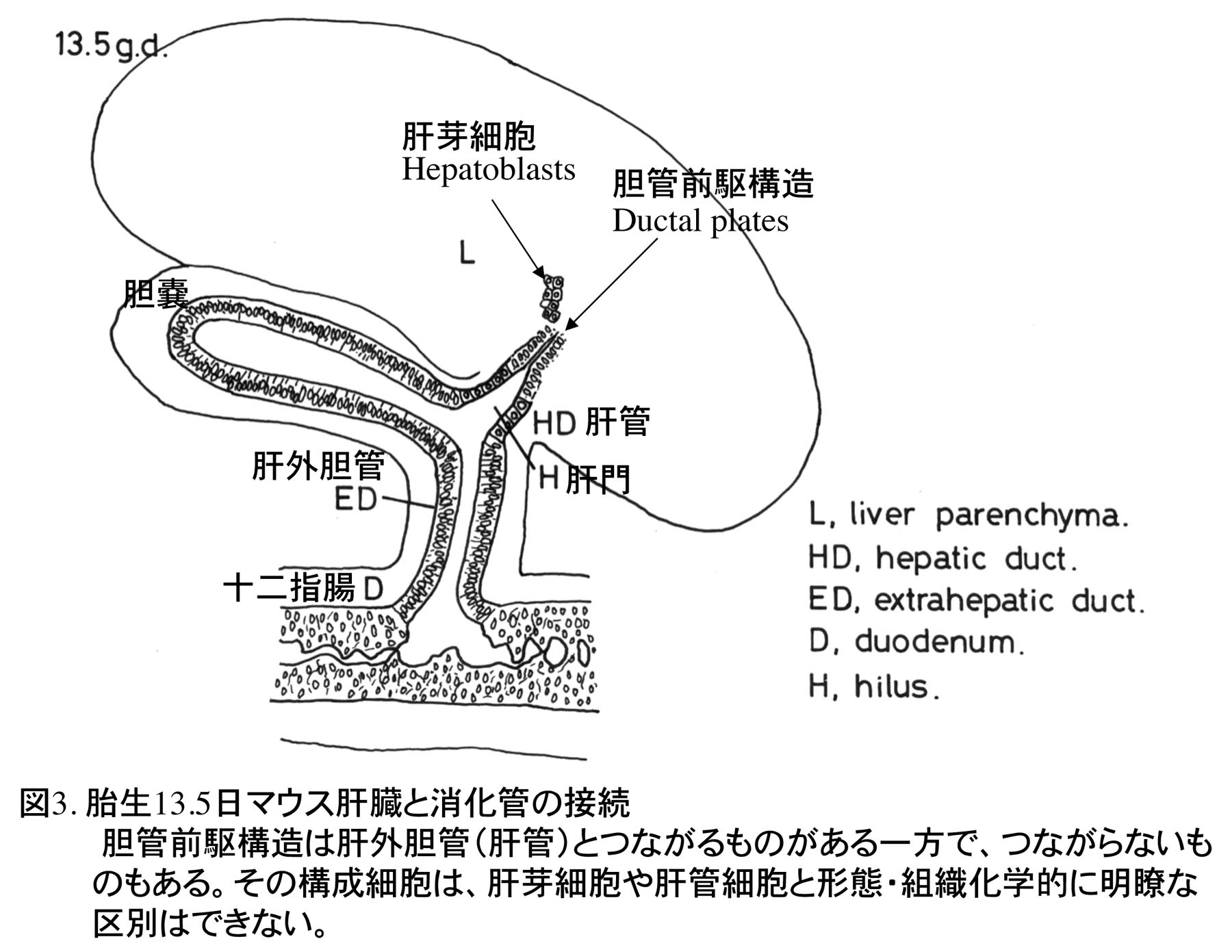
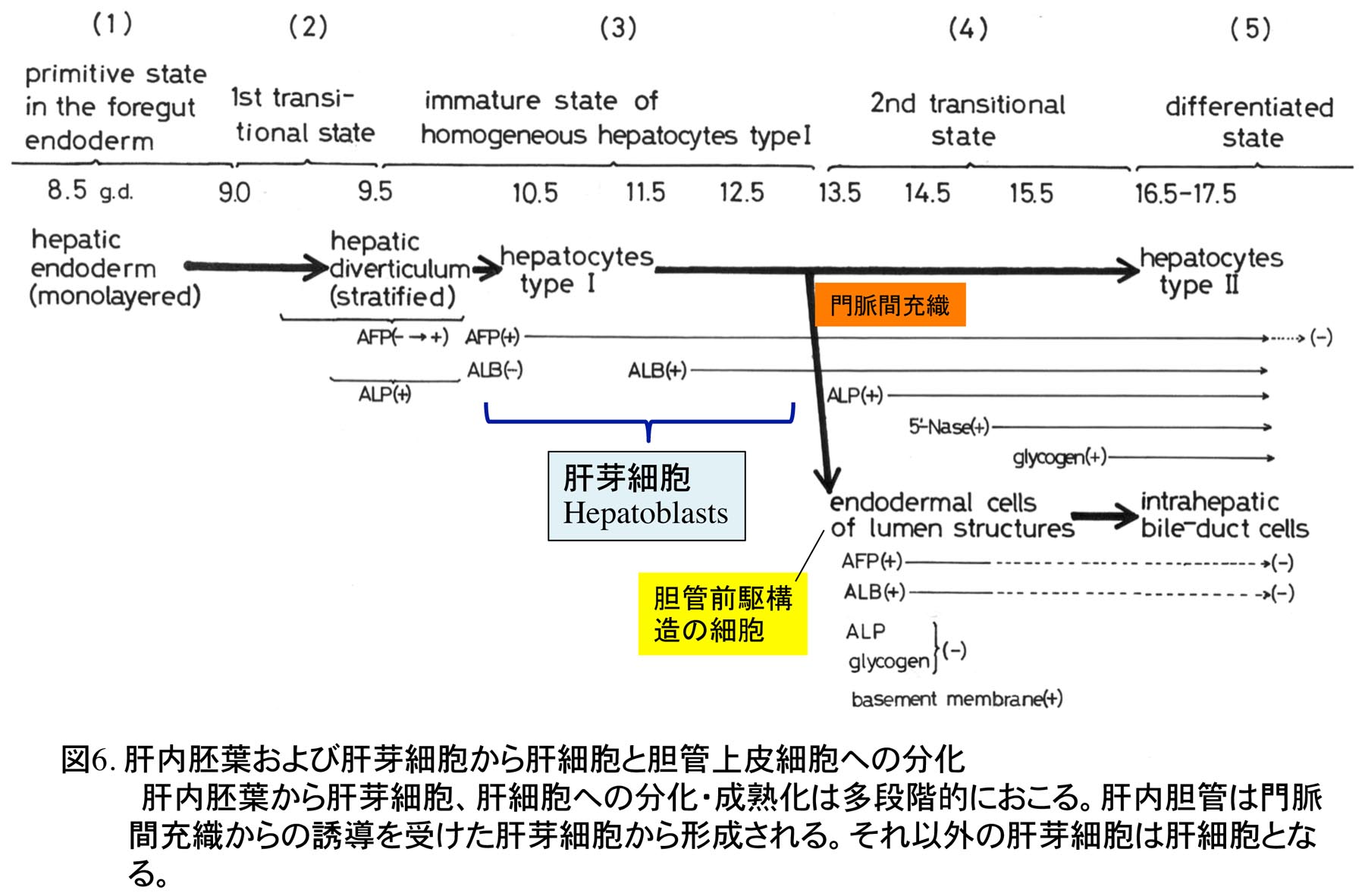
1978年に東京大学大学院理学系研究科動物学専攻に入学し、指導教授である水野丈夫先生より、マウスの肝臓発生、特に胆管発生の研究の重要性を教えられ、その研究を開始した。哺乳類胎児の肝細胞の成熟化に関する研究は当時行われていたが、胆管発生はほとんど注目されておらず、先天性胆道閉鎖症の原因等を推察する上で重要な研究であると教えられた。しかし、マウスの肝臓は胎生期に造血器官として働くため、造血細胞が多数存在し、組織学的に非常に複雑でその組織発生の理解は容易ではなかった。水野教授より、肝外および肝内胆管がどのように形成されるか、肝外胆管と肝内胆管のつながり、また肝内胆管同士のつながりについてよく観察するように指導を受け、光学顕微鏡でのマウス胎児肝臓の組織観察に加え、実体顕微鏡での肝外胆管の配向の観察とスケッチを繰り返した。マウスの肝臓原基(肝憩室)は頭部、尾部よりなり、頭部より細胞索が横中隔間充織に伸び、これが肝臓本体になり、尾部より胆嚢と肝外胆管が発生する(図1)[1]。胆嚢と肝外胆管は先に発生を開始し、発生初期の肝臓本体に肝内胆管は全く形成されていない。肝内胆管は門脈周囲に現れる胆管前駆構造より発生するが(図2)、これが未分化な肝芽細胞(私の初期の論文ではタイプI肝細胞としていた)から由来するのか、肝門に分布する肝外胆管の一つである肝管が門脈に沿って伸びることで形成されるのかが、大きな問題であった。胆管前駆構造は肝芽細胞と肝管の細胞両方に接続し、形態学的にも類似しており、どちらから形成されるか、組織学的には結論づけられなかった(図3)。なお胆管前駆構造に対し、私自身は当時”lumen structures”と命名したが、後に、ベルギーの病理学者Desmet教授たちがヒトの胆管前駆構造に対し”ductal plates”あるいはラットのそれに対し”pearl-like structures”と名づけた[2,3]。現在では、ラット、マウスの胆管前駆構造についても”ductal plates”が定着している。
肝内胆管の起源を探るべく、組織学的解析の次に、胎児肝細胞あるいは肝芽細胞で発現するAFPやアルブミンの免疫組織化学、毛細胆管アルカリ性フォスファターゼ活性等の酵素組織化学、透過型電子顕微鏡による微細形態の観察により肝芽細胞、胆管前駆構造と肝管の細胞の比較解析を行った[4-6]。胆管前駆構造の細胞は、AFP陽性、アルブミン陽性で、肝芽細胞に発現パターンが類似すると同時に、肝管細胞も両マーカーについて陽性のものがあり、肝芽細胞と肝管細胞の中間型と結論づけられた。透過型電子顕微鏡の観察では、胆管前駆構造の細胞は、肝管の細胞同様、基底膜をもっていた。肝芽細胞は基底膜をもたない。また、門脈周囲に胆管前駆構造が形成され始める時期に、門脈周囲以外の肝細胞で毛細胆管アルカリ性フォスファターゼ活性の発現が始まり、この酵素活性が陰性の胆管前駆体の細胞とは機能的に異なり始めていることが明らかとなった。これらの結果は、肝内胆管形成と肝細胞の成熟化がほぼ同時に進行していることを示唆している。いずれにせよ、以上の解析からも肝内胆管の起源を結論づけることはできなかった。
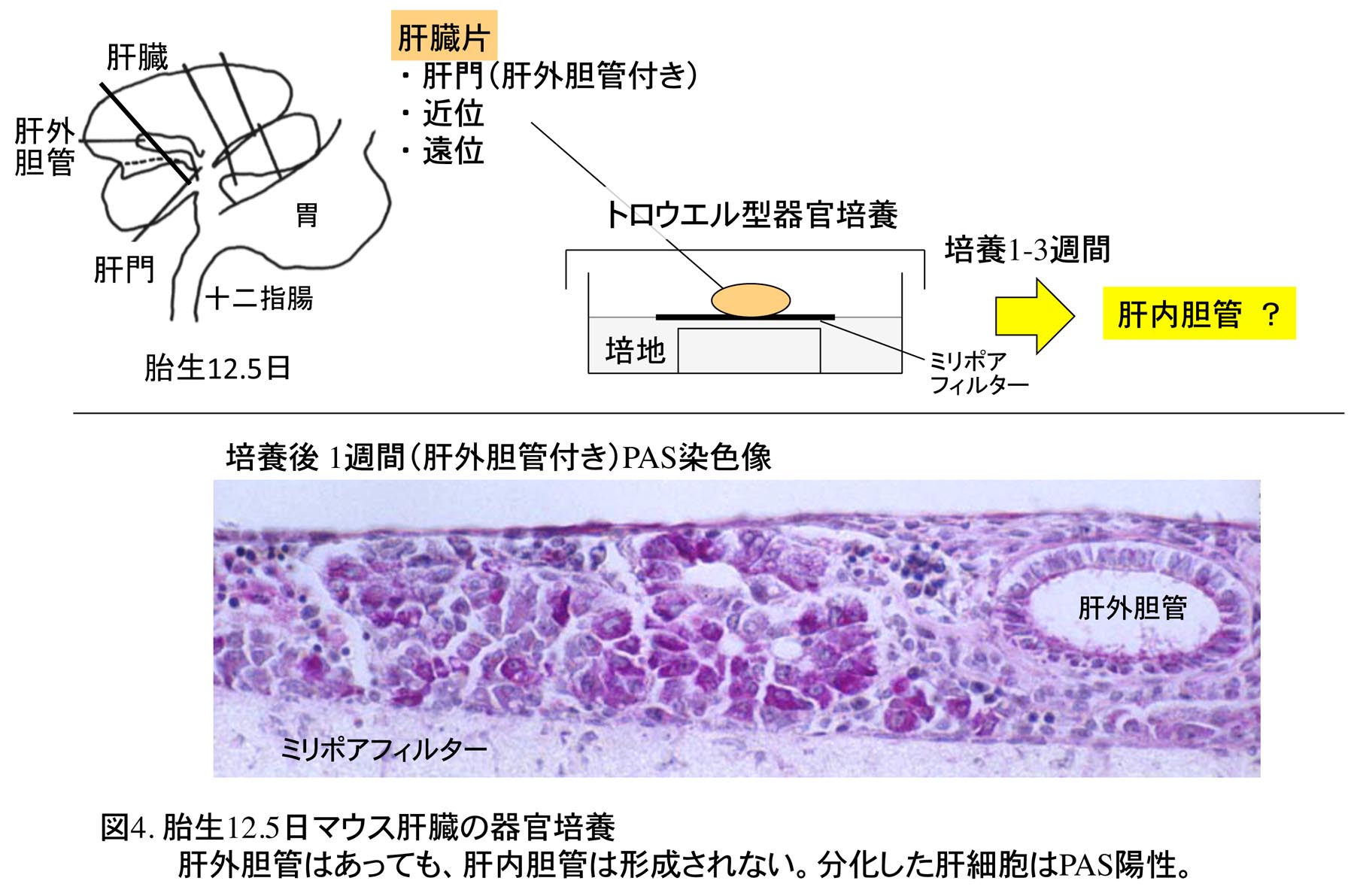
当時の仮説の一つとして、肝内胆管は肝芽細胞から門脈組織からの誘導を受け分化するとされていて、もしそうならば、肝内胆管が形成される前の肝臓片(門脈を含む)をインビトロで器官培養すると、肝内胆管が形成されるのではないかとの考えのもと、大学院時代に培地や共培養する組織を変えて培養実験を本当に何度も行った(図4)。しかし、結果として、肝内胆管を作ることはできなかった。肝外胆管をつけたまま肝門部に近い肝臓片で太い門脈が含まれているものを培養しても肝内胆管はできない[7]。肝内胆管の形成をインビトロで進めるには特別な何かが必要なようである。この方向での培養実験はなかなか成功の見込みがないために、博士課程では1年間研究テーマをラットの前立腺の発生に変更した。
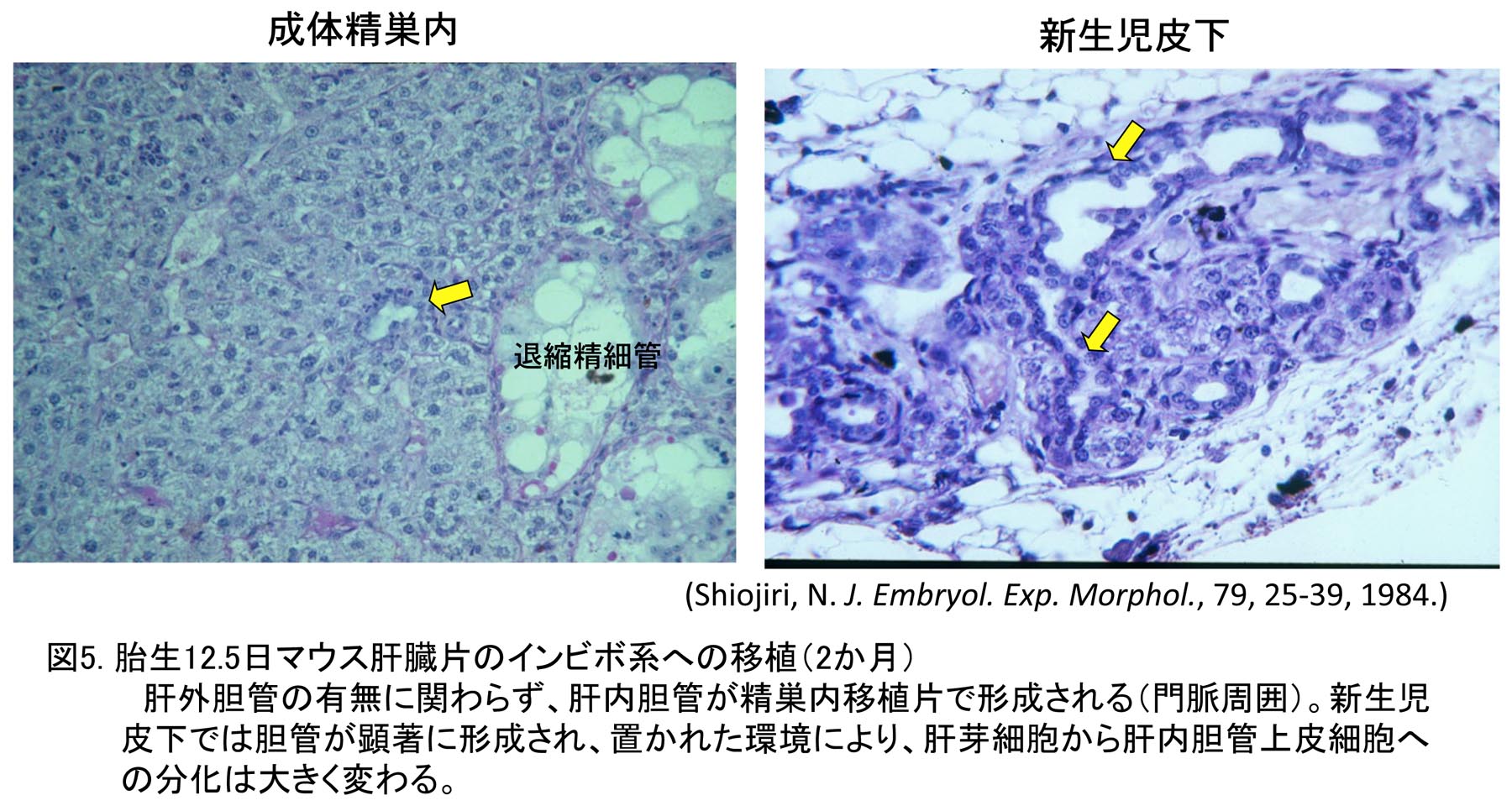
研究室の先輩より、インビトロでの肝組織の培養は限界があるので、インビボへの移植が有効ではないかとアドバイスを受け、胎生期マウス肝臓片のニワトリ胚漿尿膜への移植実験をまず行なった。しかし、この移植実験では、類洞の発生を含め肝臓の組織形成がうまく進まなかった。そこで、同じ鳥類であるウズラ胚の組織はニワトリ胚漿尿膜上で組織形成が進むとされているので、ウズラ胚の肝臓発生を押さえた後、肝内胆管前駆構造が出現する前の未分化な時期の肝臓片を同様にニワトリ胚漿尿膜上に移植した。その結果、移植したウズラ肝組織において組織形成が進み、肝内胆管が形成された[8]。この結果より、鳥類胚では、肝内胆管は肝芽細胞が門脈からの誘導を受けて形成される可能性が高いと考えた。マウス胎児肝臓組織のニワトリ胚漿尿膜上への移植結果は哺乳類と鳥類という系統的な違いに由来する可能性が高い。そこで次に、マウス胎児肝臓組織をマウス体内の種々の場所に移植を行なった(移植期間2ヶ月間)。その結果、精巣内の環境で最も組織形成が進むとともに肝内胆管が形成された(図5)。一方、新生児皮下では胆管が顕著に形成された[9]。これらのデータより、肝芽細胞は肝細胞と胆管上皮細胞の両方に分化すると結論した(図6)。しかも肝芽細胞から肝細胞と胆管上皮細胞への分化は置かれた環境に大きく影響を受け、皮下に置かれた場合その結合組織が肝芽細胞から胆管上皮細胞への分化を促進した可能性を考え、正常発生では、門脈間充織が肝芽細胞に対し誘導作用を及ぼし、肝内胆管が形成されると考えた。これらの成果により学位を取得した。
Nelson Fausto教授とoval細胞
学位取得後、静岡大学理学部生物学科に1983年4月に着任した。マウスの肝臓発生の研究は学位論文の印刷公表の問題もあり継続したが、特に胆管形成についてはインビトロでの解析が難しいことから、肝臓研究を継続すべきかあるいは新しい研究テーマに変えるべきか決めきれず、ニワトリ胚を用いて細胞接着の仕組みや、小腸の絨毛形成およびファブリキウス嚢に関する研究を試行的に進めた。当時所属した講座の片山一教授(静岡大名誉教授)は肝臓の研究を続けたらいいと言ってくれていたが、それでも学位取得後の研究テーマについて整理できなかった。そんな状況にあった1987年9月に、肝再生や肝幹細胞に関する研究を精力的に行なっていた米国ブラウン大学医学部のNelson Fausto教授より、共同研究の申し入れがあった。Fausto教授は、私が1981年の日本発生生物学会で口頭発表を行った際の英文要旨にある、精巣に移植されたマウス胎児肝臓片で、肝芽細胞が成熟肝細胞に分化するとともに肝内胆管を形成するという内容に強く関心を持ち、肝幹細胞の一つと考えられていたoval細胞の分化能について発生学的な観点から共同研究をしないかと提案をしてくれた。片山先生にいい機会だから勉強してこいと背中を強く押してもらい、1988年から1989年にかけブラウン大学に10ヶ月滞在し、oval細胞が肝芽細胞と同様に本当に未分化な細胞であるか、そのcharacterizationと分化誘導を行うことになった。
ブラウン大学での動物材料はラットで、その肝臓の発生についてAFPやアルブミンのインサイチュハイブリダイゼーション、サイトケラチンの免疫組織化学的解析等を行い、マウス胎児で得た結論と同様の結論を得た[10]。oval細胞のcharacterizationとしては、胆管上皮細胞にコミットした細胞であると考えた。発生学的には出生前の胆管前駆構造の細胞あるいは胆管上皮細胞に相当する。また、Fausto教授の研究室の大学院生とともに、ガラクトサミン投与により肝障害をおこしたラットで肝臓に出現するoval細胞での遺伝子発現や細胞増殖パターンを解析し、肝細胞に増殖が認められる前に胆管上皮細胞で顕著に増殖が起こることから、oval細胞は胆管上皮細胞由来と結論づけた[11]。oval細胞はAFP mRNAを発現する。さらに、oval 細胞の分化誘導については、Fausto教授より、ニワトリ胚の発生過程で肝臓が形成されるには心臓中胚葉が内胚葉に対し誘導することが不可欠であるという論文報告があるので、oval細胞の細胞株をラット胎児の心臓中胚葉と共培養すると肝細胞を誘導できるのではないかと提案を受け、その実験を実施した。しかし、残念ながら結果として細胞株から肝細胞を得ることはできなかった。これらの実験に示されるように、Fausto教授は実験病理学が専門ではあるが、発生学等の情報を精力的に集めるなど、視野が広く、柔軟な発想を有する勉強家であった。
Fausto教授に招待され、ブラウン大学で研究を行っていた時、当時の米国の肝臓研究者が東大の紀要に私が発表した論文[1]のコピーを持っていて、注目してくれていたことを知り、大変驚くとともに、肝臓発生の組織学については自分自身の強みであると実感した。またFausto教授により、米国やカナダの肝臓研究者、Norman Marceau教授、Kenneth Zaret教授、Douglas Hixson教授、Stewart Sell教授等を紹介されたことは、Fausto教授自身やその研究室の仲間とのつながりを含め、共同研究の実施、国際シンポジウムへの招待や論文査読など、その後の私のキャリアの中で大きな財産となった。研究面にとどまらず、互いを理解し合える友人・仲間が国際的にいてくれることは大変幸せなことである(図7)。

肝臓研究の再出発
米国留学後、大学院時代およびブラウン大学での研究成果、そして国外の研究者のコミュニテイからの認知も踏まえ、Fausto教授との共同研究はもちろん、マウスの肝臓発生に関する研究を継続することに決めた。Fausto教授からはいつ研究室に来てもいいと言ってくれたが、その共同研究を静岡大学でそのまま行うことは予算的にできず、静岡大学では学生が参加してくれるメリットを活かしつつ、肝臓に関わる特色のある研究を行うべきと考えた。それまでの経験から、組織形態学、微小の組織を扱う技術と培養系、そしてユニークな動物モデルの活用などを柱として、肝臓発生・再生の仕組みの解明を目指し再出発を行った。ユニークな動物モデルについては、マウスで遺伝子改変技術がすでに開発されていたが、地方国立大学でその技術を導入することは相当きびしいと考え、まずはすでに発表されている肝臓異常を示す変異体を探した。静岡県立大学の中村明教授より肝臓の初期発生に異常を来すとされるウズラ黒色初毛致死突然変異体(BH)を教えていただき、それを解析することにした。結果、この変異ウズラでは、ホモ型肝臓で肝細胞の壊死が高頻度で起こるものの肝臓の初期形成過程に異常が起こるわけではなかった。しかし、羽毛芽の紋様ならびに肺や皮下の血管形成が大きく乱れることが明らかとなり、その研究も進めた[12,13]。また肝臓でのオルニチントランスカルバミラーゼの発現がモザイクになるspfashヘテロ型マウスを用いて、肝臓発生・胆管発生ならびに肝再生等における細胞行動の解析を行うこととした[14,15]。さらに、非常に小さいマウス肝臓原基における組織間相互作用を証明する実験などにもチャレンジし[16,17]、またさらに時期はずれるが、脊椎動物における肝臓の形態進化等についても研究を展開した[18,19]。肝臓は脊椎動物に固有の臓器であるが、肝臓がどのように形態進化したか明らかにするため、祖先的な脊椎動物である円口類ヌタウナギやスナヤツメをはじめ、200種以上の脊椎動物の肝臓構築を調べた。ハイギョ類の肝臓構築は四足類のものに近いと推察されたが異なっていたため、入手が限られる「生きている化石」シーラカンスの肝臓構築も解析した[20]。
市原明先生(当時徳島大学教授)が始められた初代培養肝細胞研究会はある意味で肝細胞研究会の前身でもあるが、私自身は初代培養を行っていなかったため研究成果をこの会で発表することはほとんどなかった(1991年の札幌での第7回初代培養肝細胞研究会においてトピック講演として肝臓における細胞系譜について話をさせていただいた)。初代培養肝細胞研究会は徳島で開催された第9回で最後となり、1994年に肝細胞研究会が新たに発足した。肝細胞研究会は「肝臓を構成する細胞に関する研究の発展向上を図ることを目的」とする会で、初代培養を扱わない肝臓の研究も発表できることになり、その第1回が、赤池敏宏先生(当時東工大教授)が会長をされて東京で開催された際には、マウスの肝臓原基の器官培養を行った結果の発表を行った。以来、肝細胞研究会には毎回出席し発表を行ってきた。肝細胞研究会は基礎と臨床の先生方が出席され、医学的観点での肝臓の見方、薬学あるいは工学的なアプローチなど、多くのことを勉強させていただいた。専門分野の異なる研究者が一堂に介し、分野横断的に肝臓の構造・生理から病態、そして治療法の確立までを議論する研究会の意義は極めて大きい。肝臓の形態進化まで議論いただけたのは私にとって幸運であったし、研究会のポテンシャルの高さを示していると思っている。研究会では若い先生方が挑戦的で、元気であり、活発なdiscussionが行われ、これらは本当に素晴らしいことである。発表を行った私の研究室の学生も暖かく鍛えていただけた。
まとめ
これまで、大学院時代の研究テーマをスタートに、遺伝子改変マウスやインビトロでの培養などを通じて肝臓の発生・再生の仕組みや肝臓構築の形態進化などに関する研究を推進してきた。特色を出したいと考えて研究を行なってきたが、振り返ると、行き当たりばったりになっていたり、また十分に解析できていないというのが実感でもある。遺伝子改変技術、ゲノム情報、細胞選別技術等、種々の解析技術が進歩し、肝臓を理解するアプローチは私が研究を開始したときに比べ、圧倒的なレベルで進化している。その結果、解き明かされた謎や答えの得られた問い・課題は少なくない。しかし、肝臓をめぐって挑戦されるべき謎や問いは依然数多く残されているのが実態でもある。
脊椎動物における肝臓の多様性と形態進化などについてさらに研究が進められたらと思っている。肝臓はなぜ脊椎動物において獲得されたのであろうか、肝臓を持たないナメクジウオやホヤで本当にどの部分が肝臓の前駆構造であるのか、真骨類は新たな肝臓構築を進化させたが、その理由は何なのかなど、肝臓の形態進化の観点からは興味深い問いが残されている。iPS細胞からの肝臓構築はすでに報告されているが、そのスケールアップも大きな課題であるし、肝内胆管が組み込まれた肝臓をインビトロで作り出すことも重要である。私の研究室で、肝芽細胞から胆管前駆構造(ductal plates)そして肝内胆管を器官培養系でひとまず作り出せるところまでは進められたが、効率よく作出できるレベルには至っていない。これは本当に残念である。さらなる研究の進展を期待したい。さらに、私たちのモザイク解析において、細胞は組織形成の過程でランダムに動くが、結果として作られる肝臓は美しい組織構築をとる。その分子基盤は何なのかも大変興味深い。肝臓の研究はまだまだ奥深いと思う。
私自身の研究の発展・深化には、色々な方との出会いが必須のものであったし、人との出会いの素晴らしさを改めて強く感じる。サポートいただいた方について何人かここで紹介したが、他にも名前をあげていない国内外の色々な方からアドバイスをいただき、また支えていただいた。この場を借りて厚くお礼を申し上げたい。また、肝細胞研究会は異分野の研究者のプラットフォームとして大変貴重なものであり、さらに発展していくことを強く願っている。
謝辞
肝細胞研究会代表世話人三高俊広先生、HP委員長汐田剛史先生、HP委員田中稔先生には、この拙文の執筆の機会を与えていただき、厚くお礼申し上げます。
文献
- Shiojiri, N. (1979) The differentiation of the hepatocytes and the intra- and extrahepatic bile duct cells in mouse embryos. J. Fac. Sci., Univ. Tokyo, IV, 14, 241-250.
- Van Eyken, P., Sciot, R., Callea, F., Van der Steen, K., Moerman, P. and Desmet, V.J. (1988) The development of the intrahepatic bile ducts in man: a keratin-immunohistochemical study. Hepatology, 8, 1586-1595.
- Van Eyken, P., Sciot, R. and Desmet, V. (1988) Intrahepatic bile duct development in the rat: a cytokeratin-immunohistochemical study. Lab. Invest., 59, 52-59.
- Shiojiri, N. (1981) Enzymo- and immunocytochemical analyses of the differentiation of liver cells in the prenatal mouse. J. Embryol. Exp. Morphol., 62, 139-152.
- Shiojiri, N. (1984) Analysis of differentiation of hepatocytes and bile duct cells in developing mouse liver by albumin immunofluorescence. Develop. Growth Differ., 26, 555-561.
- Shiojiri, N. and Mizuno, T. (1983) Analysis of the differentiation of hepatocytes and bile-duct cells in the developing mouse liver by means of electron microscopy. J. Fac. Sci., Univ. Tokyo, IV, 15, 285-293.
- Shiojiri, N. and Mizuno, T. (1993) Differentiation of functional hepatocytes and biliary epithelial cells from immature hepatocytes of the fetal mouse in vitro. Anat. Embryol., 187, 221-229.
- Shiojiri, N. and Mizuno, T. (1987) Normal development of extra- and intrahepatic bile ducts. In: Biliary Atresia (Ed. R. Ohi), pp3-9, Tokyo, Professional Postgraduate Services.
- Shiojiri, N. (1984) The origin of intrahepatic bile duct cells in the mouse. J. Embryol. Exp. Morphol., 79, 25-39.
- Shiojiri, N., Lemire, J. M. and Fausto, N. (1991) Cell lineages and oval cell progenitors in rat liver development. Cancer Res., 51, 2611-2620.
- Lemire, J. M., Shiojiri, N. and Fausto, N. (1991) Oval cell proliferation and the origin of small hepatocytes in liver injury induced by D-galactosamine. Am. J. Pathol., 139, 535-552.
- Kubota, Y., Nakamura, A., Ito, M. and Shiojiri, N. (1995) Bh (black at hatch) gene appears to cause whole-body hemorrhage in homozygous embryos of Japanese quail (Coturnix coturnix japonica). J. Exp. Zool., 271, 441-451.
- Satoh, H., Nakamura, A. and Shiojiri, N. (1997) Bh (black at hatch) gene appears to be expressed in melanocytes of feather germs in the Japanese quail (Coturnix coturnix japonica). Develop. Growth Differ., 39, 445-450.
- Shiojiri, N., Imai, H., Goto, S., Ohta, T., Ogawa, K. and Mori, M. (1997) Mosaic pattern of ornithine transcarbamylase expression in spfash mouse liver. Am. J. Pathol., 151, 413-421.
- Shiojiri, N., Sano, S., Inujima, S., Nitou, M., Kanazawa, M. and Mori, M. (2000) Quantitative analysis of cell allocation during liver development using the spfash-heterozygous female mouse. Am. J. Pathol., 156, 65-75.
- Koike, T. and Shiojiri, N. (1996) Differentiation of the mouse hepatic primordium cultured in vitro. Differentiation, 61, 35-43.
- Nitou, M., Sugiyama, Y., Ishikawa, K. and Shiojiri, N. (2002) Isolation of hepatoblasts from fetal mouse liver with magnetic beads coated with anti-E-cadherin antibodies. Exp. Cell Res., 279, 330-343.
- Umezu, A., Kametani, A., Akai, Y., Koike, T. and Shiojiri, N. (2012) Histochemical analyses of hepatic architecture of the hagfish with special attention to periportal biliary structures. Zool. Sci., 29, 450-457.
- Shiojiri, N., Kametani, H., Ota, N., Akai, Y., Fukuchi, T., Abo, T., Tanaka, S., Sekiguchi, J., Matsubara, S. and Kawakami, H. (2018) Phylogenetic analyses of the hepatic architecture in vertebrates. J. Anat., 232, 200-213.
- Shiojiri, N., Tanaka, S. and Kawakami, H. (2019) The hepatic architecture of the coelacanth differs from that of the lungfish in portal triad formation. Okajimas Folia Anat. Jpn., 96, 1-11.

